
DOI: 10.5281/zenodo.20692218 · UDC: 617.7-007.681-092:617.749
Obiective. Glaucomul primar cu unghi deschis (GPUD) este definit ca o neuropatie optică cauzată de presiunea intraoculară ridicată; cu toate acestea, cercetări tot mai numeroase sugerează că modificările metabolice și biochimice joacă un rol important în patogeneza bolii. Scopul acestui reviu este de a rezuma cele mai recente date proteomice, metabolomice și imunologice privind modificările biochimice în GPUD și de a evalua modul în care aceste modificări afectează compoziția umorii apoase.
Metode. A fost realizată o revizuire cuprinzătoare a literaturii de specialitate utilizând bazele de date PubMed, HINARI și Google Scholar. S-a pus accent pe investigațiile care utilizează profilarea proteică, metabolică și a stresului oxidativ, au fost examinate publicațiile de cercetare originale și reviurile sistematice care cercetează compoziția umorii apoase la pacienții cu GPUD. Semnalizarea receptorilor nucleari, căile metabolice, mediatorii inflamatori și markerii stresului oxidativ au fost evaluați și integrați în mod extensiv.
Rezultate. S-a demonstrat că GPUD prezintă modificări consistente în compoziția umorii apoase, inclusiv niveluri crescute de citokine proinflamatorii și componente ale complementului, dereglarea metabolismului energetic și lipidic, scăderea capacității antioxidante și creșterea indicatorilor de stres oxidativ. Analizele proteice și metabolice au demonstrat activarea căilor imunologice și inflamatorii, în timp ce enzimele glicolitice și proteinele mitocondriale sunt suprimate. Mai mult, s-a constatat că dezechilibrul metabolic și inflamația persistentă de grad scăzut în camera anterioară sunt legate de dereglarea căilor de semnalizare LXR/RXR și FXR/RXR.
Concluzii. GPUD este legat de un profil biochimic unic în umoarea apoasă, care reflectă interacțiunea dintre activarea imunologică, disfuncția metabolică și stresul oxidativ. Aceste modificări pot contribui în mod activ la disfuncționalitatea rețelei trabeculare și la progresia bolii. Profilarea bazată pe compoziția umorii apoase ar avea potențialul de a găsi biomarkeri semnificativi din punct de vedere clinic și de a crea planuri de tratament personalizate, bazate pe mecanisme.
Objectives. Primary open-angle glaucoma (POAG) is defined as an optic neuropathy due to high intraocular pressure; however, growing research suggests that metabolic and biochemical changes play an important role in disease pathogenesis. The purpose of this review is to summarize the most recent proteomic, metabolomic and immunologic data on biochemical changes in POAG and assess how these changes affect the aqueous humor’s composition.
Methods. A comprehensive literature review was conducted using PubMed, HINARI, Google Scholar databases. With a focus on investigations using proteomic, metabolomic and oxidative stress profiling, original research publications and systematic reviews examining the composition of aqueous humor in POAG patients were examined. Nuclear receptor signaling, metabolic pathways, inflammatory mediators and oxidative stress markers were all extensively assessed and integrated.
Results. POAG was shown to have consistent changes in the composition of aqueous humor, including raised levels of pro-inflammatory cytokines and complement components, dysregulation of energy and lipid metabolism, decreased antioxidant capacity, and increased oxidative stress indicators. Proteomic and metabolomic analyses showed that immunological and inflammatory pathways were activated, while glycolytic enzymes and mitochondrial proteins were suppressed. Furthermore, metabolic imbalance and persistent low-grade inflammation in the anterior chamber were found to be linked by the dysregulation of the LXR/RXR and FXR/RXR signaling pathways.
Conclusions. POAG is linked to a unique biochemical profile in aqueous humor that reflects the interaction of immunological activation, metabolic dysfunction, and oxidative stress. These changes may actively contribute to trabecular meshwork malfunction and disease progression. Beyond lowering intraocular pressure, aqueous humor-based multi-omics profiling has the potential to find clinically significant biomarkers and create personalized, mechanism-based treatment plans.
Una dintre cele mai frecvente cauze ale orbirii ireversibile în lume rămâne a fi glaucomul, ocupând locul doi. Acesta reprezintă o patologie neurodegenerativă, caracterizată prin afectarea nervului optic și pierderea progresivă a câmpului vizual [1-3].
Teoriile tradiționale ale patogeniei glaucomului incriminează mecanismul mecanic și cel vascular în apariția maladiei. Teoria mecanică susține că presiunea intraoculară (PIO) mărită comprimă axonii celulelor retiniene ganglionare pe lamina cribroasă, inducând apariția modificărilor glaucomatoase la nivelul papilei nervului optic [4]. Astfel, PIO este considerată un factor de risc major în dezvoltarea glaucomului, fiind estimat că la fiecare majorare a PIO cu 1 mmHg, există o creștere cu 10% a riscului relativ de a dezvolta glaucom primar cu unghi deschis (GPUD) [5].
Teoria vasculară se axează pe scăderea aportului de nutrienți și oxigen către nervul optic din cauza unei presiuni de perfuzie scăzute la nivel ocular, vasospasmului, care pot apărea în apneea de somn, migrenă, boli cardiovasculare, etc [6, 7]. PIO mărită are și aici un rol important, în special din cauza că poate crește rezistența fluxului sanguin la nivelul discului nervului optic, ducând la o reducere a aportului de sânge [8].
Totuși, aceste teorii au și anumite limitări, în special teoria mecanică nu poate explica patogenia glaucomului cu presiune normală, progresia patologiei în pofida diminuării PIO. Iar cea vasculară este determinată de dificultatea precizării dacă modificările vasculare preced neurodegenerarea, dificultăți în măsurarea fluxului sanguin, lipsa studiilor prospective și pe termen lung, etc [9].
Astăzi, un număr mare de studii indică faptul că modificările biochimice din interiorul ochiului sunt esențiale în inițierea și progresia bolii, mai degrabă decât a fi o afecțiune legată de presiune [10]. Aceste schimbări sunt deosebit de pronunțate în umoarea apoasă (UA), lichidul transparent din camera anterioară care are rol de nutriție a corneei și cristalinului, structuri oculare avasculare, precum și în menținerea PIO [11-13]. Astăzi, UA este considerată un biomarker important pentru sănătatea oculară, iar componența sa oferă o perspectivă asupra stării biochimice a camerei anterioare. Astfel, s-a constatat că modificările compoziției UA pot perturba homeostazia celulară, ducând la stres oxidativ, inflamație și apoptoză neuronală [14]. În glaucom, există abateri semnificative de la profilul normal al UA, inclusiv niveluri alterate de proteine, lipide, metaboliți etc.
Scopul acestei analize este de a oferi o sinteză a cunoștințelor actuale privind modificările biochimice care sunt implicate în glaucom, cu accentuarea influenței acestora asupra compoziției UA. Evidențierea acestora ar permite evaluarea biomarkerilor umorii pentru diagnosticarea precoce a bolii, evoluția acesteia, precum și noi abordări de tratament în dependență de mecanismul fiziopatologic implicat.
În cadrul acestei lucrări au fost analizate sursele bibliografice moderne privind căile biochimice implicate în glaucom, în special GPUD, care influențează compoziția UA. Au fost consultate baze medicale, precum PubMed, HINARI, Google Scholar. Căutarea s-a efectuat folosind cuvinte cheie: ”compoziția umorii apoase”, ”markeri biochimici”, ”markeri pentru glaucom primar cu unghi deschis”, ”fiziopatologia glaucomului”, ”analiză proteomică”, ”analiză metabolomică”. Au fost selectate surse de literatură publicate în ultimii 10 ani, incluzând studii originale, metaanalize și articole de sinteză care au cercetat modificările biochimice asociate glaucomului primar cu unghi deschis și impactul acestora asupra compoziției UA. Studiile cu date incomplete și cele care nu respectau standardele metodologice au fost excluse.
Dezvoltarea glaucomului primar cu unghi deschis este determinată de procese biochimice complexe, precum stresul oxidativ și afectarea sistemului antioxidant, inflamația cronică și dereglarea imună, mecanismele neurodegenerative (figura 1) [15, 16]. Aceste fenomene modifică compoziția UA (profilul proteinomic, metabolomic și lipidic) și contribuie la progresia bolii.
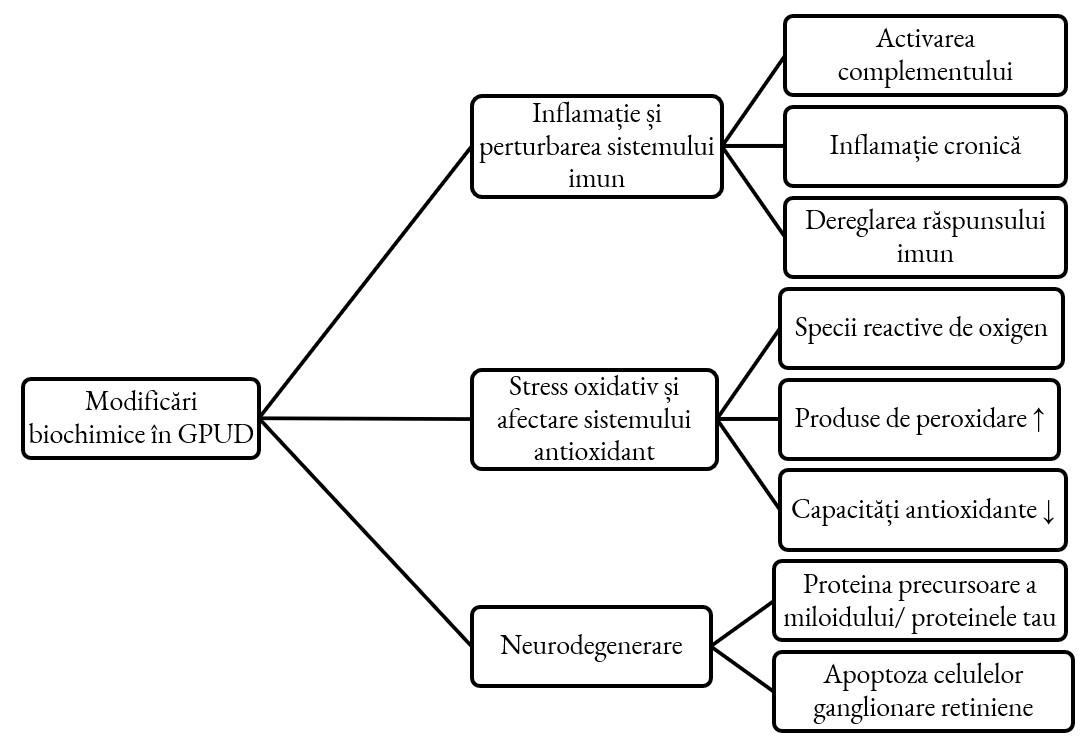
Modificări ale profilului proteic în glaucom
Analiza proteomică a UA la pacienții cu glaucom diferă semnificativ de cea a subiecților sănătoși sau a celor cu cataractă (care au servit drept grup de control). Într-un studiu vast de spectrometrie de masă, au fost identificate peste 1000 de proteine unice, cu o rată de detectare falsă mai mică de 1% [17]. În special, au fost evidențiate proteine asociate cu inflamația și răspunsul imun. Aceste procese au loc atât la nivel de rețea trabeculară, cât și la nivel de celule ganglionare retiniene (RGC).
Dintre procesele care contribuie la inflamația cronică și perturbarea sistemului imun este menționată dereglarea cascadei complementului, cu inducerea neurodegenerării. În patogenia GPUD a fost deja demonstrată implicarea cascadei complementului, în special faza sa terminală, aceasta ducând la modificarea compoziției UA. În condiții normale, sistemul complementului este implicat în diverse răspunsuri inflamatorii și de apărare ale gazdei, fiind responsabil de eliminarea celulelor degradate și a agenților patogeni, mecanisme dereglate la pacienții cu glaucom [18]. Cascada complementului contribuie semnificativ la dezvoltarea și progresia GPUD printr-o serie de mecanisme distincte, acționând ca un motor al neuroinflamației și leziunilor celulare. Această implicare este considerată a fi precoce, activarea complementului având loc chiar și în stadiile incipiente, afectând sinapsele și RGC înainte de apariția deteriorării majore a nervului optic. Odată activat, complementul dereglat declanșează un proces de neuroinflamație persistentă la nivelul retinei, care afectează direct funcția și supraviețuirea neuronilor. Un mecanism crucial de deteriorare este activarea căii finale a complementului, care culminează cu formarea complexului de atac al membranei (MAC sau C5b-9). Acest complex formează defecte în membranele RGC, conducând în cele din urmă la moarte celulară [19-21]. În plus se dereglează și procesul de eliminare a sinapselor ”mai slabe” mediat de C1Q, ducând la eliminarea sinapselor RGC esențiale. Astfel, componente specifice precum C1q, C3 și MAC sunt găsite în mod constant activate la pacienții glaucomatoși, subliniind rolul lor central în mecanismele de deteriorare neuronală (tabelul 1) [21].
Neurodegenerarea în GPUD este reprezentată de relația dintre nivelurile scăzute ale factorilor de reglare și cele crescute ale proteinelor active ale complementului din UA [20, 21]. Au fost determinate niveluri crescute ale unor componente cheie ale complementului în umoare și în țesuturile oculare, precum: C1Q, CFI, C9, VTN, C8 lanțul A, pe când C4B binding protein alpha (C4BPA), CFH, C5, C6 și C7 sunt detectate în cantități scăzute [21]. Modificări semnificative ale nivelurilor acestor proteine pot varia în funcție de rasă, sex sau stadiul bolii [20].
Componentele C5, C6, C7 și C9 sunt esențiale pentru calea terminală, care se finalizează cu formarea MAC. Activarea proteolitică a C5 este un moment crucial, deoarece produce C5A (anafilatoxină puternic proinflamatorie) și C5B, care încep să formeze secvențial componentele C6, C7, C8 și C9. Deși C5B este cel mai frecvent identificat (prezent în caz de leziuni la nivel de retină), nivelurile componenților C6, C8G sunt frecvent detectate în cantități scăzute în UA a pacienților cu glaucom. Acest lucru indică utilizarea lor în procesul de formare MAC [18, 20, 22, 23]. Factori de reglare precum factorul H (CFH), factorul I (CFI) și vitronectina (VTN) protejează celulele rețelei trabeculare și RGC de atacul complementului prin acțiunea lor. Nivelurile scăzute în umoare la pacienții glaucomatoși subliniază un eșec de reglare, care permite complementului să rămână activ și să perpetueze neuroinflamația cronică [20]. O activare cronică și necontrolată a cascadei complementului este rezultată din balanța formată (↑ activare vs ↓ reglare), care produce fragmente proinflamatoare (C3A, C5A) și complexe terminale litice (MAC/C5B-9) [20, 22, 23].
| Componentul complementului | Nivelul relativ în GPUD vs control | Funcție | Calea principală implicată |
|---|---|---|---|
| C3 | Crescut sau modificat | Componentă centrală a tuturor căilor, scindată în C3A (anafilatoxină) și C3B (opsonină) | Clasic, alternativ, lectină |
| C4A, C4B | Crescut în unele cohorte (ex. afro-americani) | Componentă timpurie | Clasic, lectină |
| F2 (protrombină) | Crescut | Complement reglator/ factor de coagulare | Regulator |
| C6 | Scăzut | Componentă esențială pentru formare MAC | Calea terminală |
| C9 | Scăzut | Componentă finală, esențială pentru formare MAC | Calea terminală |
| CFH (complement factorul H) | Scăzut | Regulator al căii alternative, protejează celulele proprii | Alternativă (reglator) |
Prin urmare, pierderea funcției de protecție a regulatorilor împreună cu abundența componentelor active face UA un mediu neurotoxic, care provoacă inflamație și moartea progresivă a RGC, eveniment patogenetic specific GPUD [22].
Niveluri crescute de citokine proinflamatorii, precum IL-8, IL-12, IL-15, IFN-γ, MCP-1 și IP-10, au fost detectate în mod constant în UA a pacienților cu GPUD (tabelul 2). Aceste proteine indică la fel o stare de inflamație cronică de grad scăzut în camera anterioară [24-27].
| Citokinele | Nivelurile în UA | Referința |
|---|---|---|
IL-5 IL-12 IL-15 IFN-γ |
Creșteri semnificative pentru markeri pro-inflamatori (p<0,05) | Burgos-Blasco B. et al., 2020 [24], Chua J. et al., 2012 [25] |
| IL-8 | Niveluri semnificative mai ridicate la pacienții cu GPUD față de grupul de control (p<0,001) | Kuchtey J., Rezaei K.A. et al., 2010 [26], Yoichi Kadoh et al., 2025 [27] |
IP-10 MCP-1 MIP-1β |
Niveluri semnificative mai ridicate față de grupul de control (p<0.05) | Kokubun T., Tsuda S. et al., 2017 [28] |
| IL-10 | Niveluri semnificativ mai crescute la pacienții cu GPUD față de grupul de control (p<0,05) | Chua J. et al., 2012 [25], Azza D. et al., 2023 [29] |
| Niveluri semnificativ mai scăzute la pacienții cu GPUD față de grupul de control (p<0,001) | Kuchtey J. et al., 2010 [26], Kokubun T., Tsuda S. et al., 2017 [28] |
Mai multe studii au demonstrat că UA provenită de la pacienții cu GPUD prezintă o creștere semnificativă a citokinelor proinflamatorii precum IL-5, IL-12, IL-15 și INF-γ (interferon-gamma), susținând ideea că inflamația cronică de intensitate redusă este o componentă a fiziopatologiei glaucomatoase [24, 28]. Niveluri ridicate de IL-12 și IFN-γ sugerează o activitate imunitară susținută de tip Th1 (limfocite T helper tip 1), care pot declanșa stresul oxidativ mediat de oxidul nitric și remodelarea matricei extracelulare în rețeaua trabeculară [25, 30]. De asemenea, creșterea IL-15 poate contribui la supraviețuirea prelungită și activarea celulelor citotoxice și NK (natural-killer) din camera anterioară (CA), în timp ce IL-5 – deși asociată în mod clasic cu activarea eozinofilelor – ar sugera implicarea subtilă a semnalizării imune de tip 2 [24, 31]. Această activare imunitară contribuie la disfuncția rețelei trabeculare, stresul mitocondrial și creșterea progresivă a PIO [30].
IL-8 este una dintre cele mai constant crescute citokine din UA a pacienților cu GPUD, reprezentând un puternic chemoatractant neutrofil și mediator al semnalizării imune înnăscute. IL-8 contribuie la stresul rețelei trabeculare prin promovarea eliberării de specii reactive de oxigen și amplificarea remodelării țesutului local. Nivelurile crescute de această proteină proinflamatorie sunt corelate cu o apoptoză mai mare a celulelor rețelei trabeculare și o reducere a excreției UA, sugerând un rol direct în creșterea progresivă a PIO [26, 28, 32]. Mai mult decât atât, imunocolorarea cu IgG de capră, purificată pe baza afinității obținute împotriva IL-8 umane, a dus la imunoreactivitate pozitivă la nivel de nerv optic și în retina internă, nefiind determinată în alte structuri oculare. Aceste rezultate sugerează că IL-8 este produsă de țesuturi oculare și că această sursă locală poate contribui la detectarea ei în UA [26].
IP-10 (proteina 10 indusă de interferon-gamma), MCP-1 (peptidul - 1 chemoatractant al monocitelor), MIP-1β (proteina inflamatorie a macrofagelor 1 beta) sunt frecvent mărite în UA a pacienților glaucomatoși, indicând o inflamație cronică ușoară și potențiale leziuni ale celulelor ganglionare retiniene prin atragerea celulelor imune (monocite, macrofage) [24, 25]. În studiul efectuat de Taiki Kokubun și colab. a fost demonstrată corelarea semnificativă între valorile presionale cu nivelurile acestor citokine la pacienții cu glaucom primar versus pacienții cu cataractă care au servit drept lot de control. Au fost evidențiate cantități mărite de MCP-1 și MIP-1β pe măsura progresării patologiei, sugerând rolul lor în patogeneză și utilizarea lor în calitate de biomarkeri, deși nivelurile exacte ale acestora ar putea varia între studii și populații [28].
IL-10 este o citokină antiinflamatorie importantă, însă cercetările nivelului acestei proteine sunt controversate în literatura de specialitate, reflectând natura complexă și adesea contradictorie a neuroinflamației în glaucom. Multiple studii sugerează niveluri de IL-10 sunt semnificativ mai crescute în UA a pacienților cu GPUD în comparație cu cei cu cataractă. Creșterea acesteia ar putea reprezenta un mecanism compensator sau de neuroprotecție locală a țesuturilor oculare de a combate inflamația cronică și leziunile tisulare cauzate de PIO ridicată și stresul oxidativ asociat cu glaucomul [25, 29]. În același studiu realizat de Kokubun T. și colab. accentuează ratele scăzute de IL-10 la pacienții cu glaucom primar cu unghi deschis, subliniind mecanismele defecte în prevenirea apoptozei RGC [28].
De asemenea, analizele proteomice recente ale UA la pacienții cu glaucom au evidențiat o îmbogățire semnificativă a căilor de semnalizare ale receptorilor nucleari, în special activarea LXR/RXR (Receptor X Hepatic/Receptor X Retinoidic) și FXR/RXR (Receptor X Farnesoid/Receptor X Retinoid), care sunt esențiale pentru metabolismul lipidic, homeostazia colesterolului și reglarea inflamatorie în țesuturile oculare [33]. Expresia modificată a proteinelor legate de aceste căi sugerează că semnalizarea lipidică dereglată și dezechilibru metabolic pot contribui la disfuncția rețelei trabeculare și la răspunsurile inflamatorii cronice în GPUD [34]. Când sunt funcționali heterodimerii LXR/RXR, care sunt activați de oxisterolii endogeni și genele modulatoare implicate în efluxul colesterolului și răspunsurile antiinflamatorii, par să aibă un efect protector și antiinflamator în afecțiunile glaucomatoase [35]. În plus, modularea RXR a fost legată de efectele neuroprotectoare asupra RGC și reducerea răspunsului la stres. Acest lucru subliniază rolul semnalizării retinoide în fiziopatologia glaucomului [36]. În contextul biochimic al glaucomului, aceste constatări susțin ideea că activarea imunitară și neurodegenerarea sunt legate de disfuncția metabolică și căile inflamatorii reglate de lipide.
Modificări metabolice în glaucom
Analizele UA colectate de la pacienții diagnosticați cu GPUD au arătat în mod constant că glaucomul este asociat cu modificări extinse ale căilor metabolice importante (figura 2). Aceste modificări se reflectă în disfuncția bioenergetică, în remodelarea lipidelor și în metabolismul aminoacizilor, demonstrând că patologia glaucomatoasă afectează nu numai presiunea mecanică, ci și metabolismul celular fundamental [10, 37].
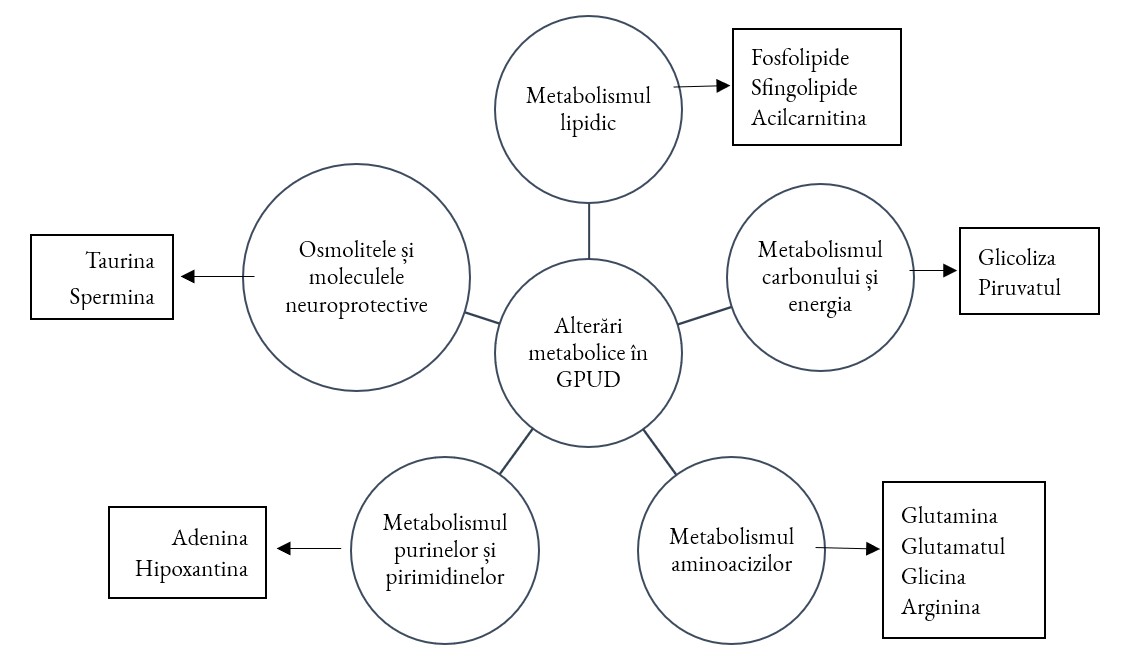
Niveluri modificate ale mai multor aminoacizi au fost raportate în mod repetat în UA a pacienților cu GPUD [10]:
Glutamina,
Glicina,
Leucina,
Histidina,
Fenilalanina,
Glutamatul,
Alanina,
Izoleucina,
Lizina,
Arginina.
Aceste modificări indică o afectare a căilor de biosinteză și degradare a aminoacizilor, inclusiv a metabolismului argininei și prolinei, glutaminei/glutamatului, glicinei și a biosintezei de aminoacil-ARNt. Aceste căi sunt direct legate de echilibrul azotului, ciclul neurotransmițătorilor, producția de energie și protecția antioxidantă. Acest proces leagă perturbarea metabolică de vulnerabilitatea RGC și disfuncția trabeculară [10].
De asemenea, metabolismul lipidic și remodelarea membranară sunt afectate în glaucom, prin dereglarea fosfatidilcolinelor, sfingomielinelor, acilcarnitinelor cu lanț scurt și alți derivați lipidici. Aceste modificări pot contribui la stresul, inflamația și reglarea osmotică afectată din CA și subliniază compoziția modificată a membranei, oxidarea acizilor grași [37]. Oxilipinele, care sunt metaboliți ai acizilor arahidonic și linoleic, subliniază rolul mediatorilor lipidici inflamatori în GPUD [38].
S-a demonstrat că metaboliții care participă la glicoliză, metabolismul piruvatului, calea pentozofosfatului și lanțul de transport al electroliților sunt alterate în UA a pacienților cu glaucom. Acest fapt indică o modificare a metabolismului carbonului și a bioenergeticii mitocondriale în condiții glaucomatoase. Acesta face ca producția de ATP (adenozin trifosfat) să scadă și să crească stresul oxidativ. Întreținerea și supraviețuirea celulară în rețelele trabeculare și neuronii retinei pot fi compromise dacă există o scădere a compușilor legați de energie [39].
Metabolismul purinelor și al pirimidinelor suferă schimbări semnificative, inclusiv modificări ale nucleotidelor (adenina și hipoxantina), fapt demonstrat în mai multe studii. Acest lucru indică o perturbare a metabolismului acidului nucleic și a echilibrului energetic, ceea ce poate afecta replicarea celulară, repararea și răspunsul la stres [40]. Compuși precum taurina și spermina, care funcționează ca osmoprotectanți și agenți neuroprotectori, sunt prezenți în cantități mici în UA în caz de GPUD. În lipsa acestora este afectat mecanismul de protecție împotriva leziunilor oxidative [37].
Combinația acestor modificări metabolice indică faptul că GPUD implică o schimbare sistemică a homeostaziei. Multitudinea de procese care perturbă producerea de energie, stresul oxidativ și semnalizarea inflamatorie interacționează pentru a afecta fluxul din rețeaua trabeculară și a accelera pierderea celulelor ganglionare retiniene. Aceste date susțin ipoteza că reprogramarea metabolică este un factor determinant al fiziopatologiei glaucomatoase și nu doar o consecință a bolii.
Stresul oxidativ
Un mecanism biochimic fundamental responsabil de patogeneză a glaucomului primar cu unghi deschis este stresul oxidativ. Umoarea apoasă din CA este expusă în mod continuu la specii reactive de oxigen (ROS) produse în țesuturile oculare în urma acțiunii radiației ultraviolete, de procese metabolice și disfuncții mitocondriale. În condiții fiziologice normale sistemele de apărare antioxidantă, care includ superoxid dismutaza, glutation peroxidaza, catalaza și antioxidanții neenzimatici (vitamina C și E) mențin homeostazia redox. Acest echilibru este perturbat în glaucom, ceea ce duce la acumulare de ROS în exces și la leziuni oxidative ulterioare (tabelul 3) [41-43].
| Marker | Nivelul în GPUD vs control | Efect reprezentativ | Valoare p | Note asupra studiului | Referința |
|---|---|---|---|---|---|
| Malondialdehida (MDA) | ↑ | Creșterea nivelului de peroxidare lipidică | p < 0,05 | Măsurători observaționale ale UA în studii caz-control | Benois d’Azy et al., 2016 [44] |
| 8-hidroxi-2-deoxiguanosina | ↑ | Markeri crescuți ai leziunilor oxidative ale ADN-ului detectat în UA la pacienți cu GPUD | p < 0,05 | Determinat prin ELISA sau HPLC din UA | Benois d’Azy et al., 2016 [44] |
| Superoxid dismutaza (SOD) | ↑ | Activitate crescută, interpretată ca răspuns compensator la stresul oxidativ | p < 0,001 | Analize enzimatice sau proteomică | Goyal A. et al., 2014 [45], Takayanagi Y. et al., 2020 [46] |
| Glutation peroxidaza (GPx) (activitatea) | ↓ | Activitate Gpx scăzută sau niveluri reduse ale enzimelor legate de glutation în multe studii privind UA | p < 0,05 | Teste de activitate enzimatică, susține apărarea antioxidantă epuizată | Benois d’Azy et al., 2016 [44] |
| Glutation S-transferaza (GST) | ↓ | Expresia/ activitatea redusă a enzimelor din familia GST | p < 0,05 | Identificat prin teste specifice și prin profilarea proteomică a UA | Liu A et al., 2021 [47] |
| Sintaza inducibila de oxid nitric (iNOS/NOS2) markeri nitrozativi | ↑ | Expresia crescută a NOS2 și creșterea markerilor de stres nitrozativ raportate în UA | p < 0,05 | Identificarea proteomică și câteva teste biochimice, sugerează o componentă nitrozativă a dezechilibrului redox | Canto A. et al., 2019 [48] |
| Capacitatea antioxidantă totală (TAC) | ↓ | Capacitatea antioxidantă redusă în UA | p < 0,05 | Studii caz-control utilizând teste TAC colorimetrice/ chemiluminescente |
Benois d’Azy et al., 2016 [44] |
În comparație cu grupul de control, alcătuit din pacienții cu cataractă de aceeași vârstă, pacienții cu GPUD prezintă niveluri mai mari ai markerilor leziunilor oxidative (malondialdehida (MDA) – un produs major al peroxidării lipidelor), produselor oxidative ale ADN-ului (8-hidroxi-2-deoxiguanosina). Acest fapt reflectă prezența peroxidării lipidice crescute și a leziunilor acidului nucleic din țesuturile oculare expuse la generarea continuă de ROS [44, 45]. În același timp capacitatea totală antioxidantă este redusă în CA a pacienților cu glaucom, ceea ce indică faptul că sistemul de apărare antioxidantă este epuizat sau suprasolicitat. Anumiți antioxidanți enzimatici, precum glutation peroxidaza și superoxid dismutaza prezintă activități modificate. Unele studii indică o creștere a activității enzimelor ca răspuns compensator la nivelurile ridicate de ROS, în timp ce altele semnalează niveluri scăzute de expresie în UA, ceea ce sugerează o afectare structurală a sistemului antioxidant în rețeaua trabeculară și țesuturile ciliare [42, 45].
Studiile bazate pe determinarea compoziției proteinemice au identificat niveluri scăzute ale enzimelor antioxidante cheie (glutation tranferaza), precum și o expresie crescută a sintazei inductibile de oxid nitric. Aceste procese duc la stres nitrosativ și la formare de specii toxice de azot, exacerbând leziunile oxidative [48, 49]. Aceste modificări oxidative ale UA sunt asociate cu disfuncția rețelei trabeculare și vulnerabilitatea structurală, ceea ce contribuie la rezistența crescută la excreția umorii și la leziuni glaucomatose progresive.
Rezultatele sistematizate în acest reviu susțin ideea că glaucomul primar cu unghi deschis este caracterizat prin multitudine de procese biochimice complexe, care influențează și compoziția UA. La nivelul rețelei trabeculare și a celulelor ganglionare retiniene, factorii care contribuie la patogeneza glaucomului includ disfuncții metabolice, stresul oxidativ, activare imunitară și semnalizare neurodegenerativă. În afară de presiunea intraoculară crescută, există dovezi convergente din studiile proteomice, metabolice și imunologice [9].
Astfel, GPUD afectează direct și cumulativ compoziția UA. Suprimarea căilor metabolice legate de energie (glicoliza și metabolismul mitocondrial) este una dintre cele mai consistente constatări din studiile proteomice și metabolice [39]. O insuficiență bioenergetică în țesuturile oculare glaucomatoase este indicată de niveluri reduse de enzime glicolitice și intermediari, precum și metaboliți alterați ai ciclului tricarboxilic [37, 39, 40]. O astfel de compromitere metabolică poate face celulele rețelei trabeculare mai susceptibile la deteriorarea oxidativă și senescență, contribuind în cele din urmă la afectarea fluxului de lichid apos și la creșterea susținută a presiunii intraoculare. În plus, ideea că stresul metabolic se extinde dincolo de segmentul anterior și poate afecta vulnerabilitatea celulelor ganglionare retiniene este susținută de lipsa metaboliților neuroprotectori și a proteinelor de susținere neuronale.
În GPUD, afectarea metabolică este strâns legată de leziunile structurale și funcționale provocate de stresul oxidativ. Un dezechilibru continuu între generarea speciilor reactive de oxigen și neutralizarea acestora este demonstrat de nivelurile crescute ale produselor de peroxidare lipidică și ale markerilor de leziuni oxidative ale ADN-ului în UA [10, 37], însoțite de o reducere a capacității antioxidante totale și de dereglarea enzimelor antioxidante importante [44-47]. Acest dezechilibru redox are potențialul de a agrava disfuncția rețelei trabeculare prin schimbări ale citoscheletului, remodelarea matricei extracelulare și creșterea rigidității celulare [46]. Este important de menționat că stresul oxidativ afectează și căile de semnalizare ale sistemului imunitar, amplificând reacțiile inflamatorii din CA.
În acest context, reglarea citokinelor și chemokinelor proinflamatorii, cum ar fi IL-8, IL-12, IL-15, IFN-γ și mediatorii imunitari asociați, oferă dovezi suplimentare că GPUD este legat de o stare de inflamație cronică de grad scăzut [24, 28]. Citokinele menționate mai sus au capacitatea de a stimula stresul celular al rețelei trabeculare, apoptoza și remodelarea fibrotică, influențând în același timp procesele neuroinflamatorii asociate cu degenerarea celulelor ganglionare retiniene [28]. Analizele proteomice ale UA au arătat activarea simultană a componentelor cascadei complementului. Acest fapt susține și rolul mecanismelor imune înnăscute în progresia glaucomului.
Dezechilibrul căilor de semnalizare LXR/RXR și FXR/RXR poate duce la dereglare metabolică și imunitară în GPUD, deoarece acestea sunt căi ale receptorilor nucleari care controlează metabolismul lipidic, homeostazia colesterolului și suprimarea inflamatorie [33-36]. Aceste procese biochimice interdependente creează un micromediu patogen în UA, influențând leziunile glaucomatoase prin perpetuarea disfuncției trabeculare și neurodegenerării retiniene.
Per ansamblu, aceste modificări biochimice din componența UA indică că glaucomul primar cu unghi deschis este mai degrabă o patologie metabolică-inflamatorie multifactorială decât o neuropatie optică cauzată exclusiv de presiune. Determinarea componentelor UA ar putea sublinia utilitatea potențială acestora ca surse de biomarkeri pentru stadializarea bolii, monitorizarea progresiei și evaluarea răspunsului terapeutic. În plus, abordarea rezilienței metabolice, a stresului oxidativ și a modulării imune – atât separat, cât și în combinație cu strategii de reducere a PIO – poate fi o direcție promițătoare pentru viitoarea terapie a glaucomului.
Se impun cercetări viitoare pe studii longitudinale care ar clarifica legătura dintre factori și ar determina modificările biochimice timpurii care preced leziunile structurale ireversibile. În cele din urmă, utilizarea acestor abordări integrative ar putea ajuta la implementarea unui management personalizat, bazat pe compoziția UA, mecanisme specifice glaucomului primar cu unghi deschis.
Glaucomul primar cu unghi deschis este asociat cu procese biochimice distincte în umoarea apoasă, reflectând convergența stresului oxidativ, disfuncției metabolice și activării imune. Detectarea consistentă a proteinelor modificate, metaboliților, citokinelor și markerilor stresului oxidativ susțin conceptul că compoziția umorii apoase reflectă procesele patogene cheie care au loc în segmentul anterior.
Compoziția UA glaucomatoase este influențată de stres oxidativ, reprogramare metabolică, activare imunitară și semnalizarea dereglată a receptorilor nucleari. Acest lucru afectează apărarea antioxidantă, metabolismul energetic și lipidic, fiind stabilită inflamația cronică de grad scăzut și activarea complementului. Aceste procese sunt legate între ele și contribuie la disfuncția rețelei trabeculare, remodelarea matricei extracelulare și creșterea rezistenței la fluxul de ieșire. În același timp, aceste procese induc afectarea celulelor ganglionare retiniene și neurodegenerare progresivă.
Modificările biochimice ale umorii apoase susțin utilitatea lor potențială ca surse de biomarkeri relevanți din punct de vedere clinic, care ar permite stadializarea bolii, identificarea precoce și monitorizarea răspunsului terapeutic. Se impun cercetări longitudinale și integrative pentru a determina relațiile cauzale și a crea abordări personalizate bazate pe mecanismele prezente în glaucom.
Quigley HA, Broman AT. The number of people with glaucoma worldwide in 2010 and 2020. Br J Ophthalmol. 2006;90(3):262-267. doi:10.1136/bjo.2005.081224
Bagnasco L, Bagnis A, Bonzano C. Terminology and guidelines for glaucoma. 5th ed. European Glaucoma Society Terminology and Guidelines for Glaucoma, 5th Edition. Savona, Italy: PubliComm; 2020. Accessed June 11, 2026. Available from: https://eugs.org/educational_materials/6
Tham YC, Li X, Wong TY, Quigley HA, Aung T, Cheng CY. Global prevalence of glaucoma and projections of glaucoma burden through 2040: a systematic review and meta-analysis. Ophthalmology. 2014;121(11):2081-2090. doi:10.1016/j.ophtha.2014.05.013
Ahmad SS. The mechanical theory of glaucoma in terms of prelaminar, laminar, and postlaminar factors. Taiwan J Ophthalmol. 2023;14(3):376-386. Published 2023 Dec 21. doi:10.4103/tjo.TJO-D-23-00103
Weinreb RN. IOP and the risk of progression to glaucoma. Graefes Arch Clin Exp Ophthalmol. 2005;243(6):511-512. doi:10.1007/s00417-005-1195-9
Chan KKW, Tang F, Tham CCY, Young AL, Cheung CY. Retinal vasculature in glaucoma: a review. BMJ Open Ophthalmol. 2017;1(1):e000032. doi:10.1136/bmjophth-2016-000032
Enyong EN, Gurley JM, De Ieso ML, Stamer WD, Elliott MH. Caveolar and non-Caveolar Caveolin-1 in ocular homeostasis and disease. Prog Retin Eye Res. 2022;91:101094. doi:10.1016/j.preteyeres.2022.101094
Flammer J, Orgül S, Costa VP, et al. The impact of ocular blood flow in glaucoma. Prog Retin Eye Res. 2002;21(4):359-393. doi:10.1016/s1350-9462(02)00008-3
Ahmad SS. Controversies in the vascular theory of glaucomatous optic nerve degeneration. Taiwan J Ophthalmol. 2016;6(4):182-186. doi:10.1016/j.tjo.2016.05.009
Tang Y, Shah S, Cho KS, Sun X, Chen DF. Metabolomics in Primary Open Angle Glaucoma: A Systematic Review and Meta-Analysis. Front Neurosci. 2022;16:835736. Published 2022 May 12. doi:10.3389/fnins.2022.835736
Benos DJ, Simon SA, eds. The eye’s Aqueous Humor. Current Topics in Membranes, Volume 62 Series Editors. 2nd ed. San Diego, CA: Elsevier Inc; 2008.
Macknight AD, McLaughlin CW, Peart D, Purves RD, Carré DA, Civan MM. Formation of the aqueous humor. Clin Exp Pharmacol Physiol. 2000;27(1-2):100-106. doi:10.1046/j.1440-1681.2000.03208.x
Iacubitchii M, Bendelic E, Alsaliem S. Aqueous humor’s biochemical composition in ocular pathologies. Mold Med J. 2019;62(2):38–43.
Böhm EW, Buonfiglio F, Voigt AM, et al. Oxidative stress in the eye and its role in the pathophysiology of ocular diseases. Redox Biol. 2023;68:102967. doi:10.1016/j.redox.2023.102967
Baudouin C, Kolko M, Melik-Parsadaniantz S, Messmer EM. Inflammation in Glaucoma: From the back to the front of the eye, and beyond. Prog Retin Eye Res. 2021;83:100916. doi:10.1016/j.preteyeres.2020.100916
Vernazza S, Tirendi S, Bassi AM, Traverso CE, Saccà SC. Neuroinflammation in Primary Open-Angle Glaucoma. J Clin Med. 2020;9(10):3172. Published 2020 Sep 30. doi:10.3390/jcm9103172
Adav SS, Wei J, Terence Y, Ang BCH, Yip LWL, Sze SK. Proteomic Analysis of Aqueous Humor from Primary Open Angle Glaucoma Patients on Drug Treatment Revealed Altered Complement Activation Cascade. J Proteome Res. 2018;17(7):2499-2510. doi:10.1021/acs.jproteome.8b00244
Ricklin D, Hajishengallis G, Yang K, Lambris JD. Complement: a key system for immune surveillance and homeostasis. Nat Immunol. 2010;11(9):785-797. doi:10.1038/ni.1923
Tezel G, Yang X, Luo C, et al. Oxidative stress and the regulation of complement activation in human glaucoma. Invest Ophthalmol Vis Sci. 2010;51(10):5071-5082. doi:10.1167/iovs.10-5289
Vashishtha A, Maina SW, Altman J, et al. Complement System Proteins in the Human Aqueous Humor and Their Association with Primary Open-Angle Glaucoma. J Pers Med. 2023;13(9):1400. Published 2023 Sep 19. doi:10.3390/jpm13091400
Hoppe C, Gregory-Ksander M. The Role of Complement Dysregulation in Glaucoma. Int J Mol Sci. 2024;25(4):2307. Published 2024 Feb 15. doi:10.3390/ijms25042307
Mitchell CH, Nickells R, Shestopalov V, Hoppe C, Gregory-Ksander M. The role of complement dysregulation in glaucoma. Int J Mol Sci. 2024;25(4):2307. doi:10.3390/ijms25042307
Mok JH, Park DY, Han JC. Differential protein expression and metabolite profiling in glaucoma: Insights from a multi-omics analysis. Biofactors. 2024;50(6):1220-1235. doi:10.1002/biof.2079
Burgos-Blasco B, Vidal-Villegas B, Saenz-Frances F, et al. Tear and aqueous humour cytokine profile in primary open-angle glaucoma. Acta Ophthalmol. 2020;98(6):e768-e772. doi:10.1111/aos.14374
Chua J, Vania M, Cheung CM, et al. Expression profile of inflammatory cytokines in aqueous from glaucomatous eyes. Mol Vis. 2012;18:431-438.
Kuchtey J, Rezaei KA, Jaru-Ampornpan P, Sternberg P Jr, Kuchtey RW. Multiplex cytokine analysis reveals elevated concentration of interleukin-8 in glaucomatous aqueous humor. Invest Ophthalmol Vis Sci. 2010;51(12):6441-6447. doi:10.1167/iovs.10-5216
Kadoh Y, Takayanagi Y, Sugihara K, Kaidzu S, Takai Y, Tanito M. Aqueous Humor Cytokine Profiling Reveals Distinct Roles for Serum Amyloid A, Interleukin-8, and Endothelin-1 in Pseudoexfoliation Syndrome and Glaucoma. Int J Mol Sci. 2025;26(4):1461. Published 2025 Feb 10. doi:10.3390/ijms26041461
Kokubun T, Tsuda S, Kunikata H, et al. Characteristic Profiles of Inflammatory Cytokines in the Aqueous Humor of Glaucomatous Eyes. Ocul Immunol Inflamm. 2018;26(8):1177-1188. doi:10.1080/09273948.2017.1327605
Dammak A, Sanchez Naves J, Huete-Toral F, Carracedo G. New Biomarker Combination Related to Oxidative Stress and Inflammation in Primary Open-Angle Glaucoma. Life (Basel). 2023;13(7):1455. Published 2023 Jun 27. doi:10.3390/life13071455
Vidal-Villegas B, Burgos-Blasco B, Santiago Alvarez JL, et al. Proinflammatory Cytokine Profile Differences between Primary Open-Angle and Pseudoexfoliative Glaucoma. Ophthalmic Res. 2022;65(1):111-120. doi:10.1159/000519816
Oribio-Quinto C, Burgos-Blasco B, Pérez-García P, et al. Aqueous Humor Cytokine Profile in Primary Congenital Glaucoma. J Clin Med. 2023;12(9):3142. doi:10.3390/jcm12093142
Khalef N, Labib H, Helmy H, El Hamid MA, Moemen L, Fahmy I. Levels of cytokines in the aqueous humor of eyes with primary open angle glaucoma, pseudoexfoliation glaucoma and cataract. Electron Physician. 2017;9(2):3833-3837. doi:10.19082/3833
Liu X, Liu X, Wang Y, et al. Proteome Characterization of Glaucoma Aqueous Humor. Mol Cell Proteomics. 2021;20:100117. doi:10.1016/j.mcpro.2021.100117
Sharma S, Bollinger KE, Kodeboyina SK, et al. Proteomic Alterations in Aqueous Humor From Patients With Primary Open Angle Glaucoma. Invest Ophthalmol Vis Sci. 2018;59(6):2635-2643. doi:10.1167/iovs.17-23434
Ju HH, Lee J, Kim SK, et al. Physiological activation of liver X receptor provides protection against ocular inflammation in uveitic glaucoma. Biochim Biophys Acta Mol Basis Dis. 2025;1871(1):167573. doi:10.1016/j.bbadis.2024.167573
Sharma S, Shen T, Chitranshi N, et al. Retinoid X Receptor: Cellular and Biochemical Roles of Nuclear Receptor with a Focus on Neuropathological Involvement. Mol Neurobiol. 2022;59(4):2027-2050. doi:10.1007/s12035-021-02709-y
Buisset A, Gohier P, Leruez S, et al. Metabolomic Profiling of Aqueous Humor in Glaucoma Points to Taurine and Spermine Deficiency: Findings from the Eye-D Study. J Proteome Res. 2019;18(3):1307-1315. doi:10.1021/acs.jproteome.8b00915
Xu J, Zhou K, Fu C, et al. Oxylipins in Aqueous Humor of Primary Open-Angle Glaucoma Patients. Biomolecules. 2024;14(9):1127. doi:10.3390/biom14091127
Monu M, Kumar B, Asfiya R, et al. Metabolomic Profiling of Aqueous Humor From Glaucoma Patients Identifies Metabolites With Anti-Inflammatory and Neuroprotective Potential in Mice. Invest Ophthalmol Vis Sci. 2025;66(5):28. doi:10.1167/iovs.66.5.28
Pulukool SK, Bhagavatham SKS, Kannan V, et al. Elevated dimethylarginine, ATP, cytokines, metabolic remodeling involving tryptophan metabolism and potential microglial inflammation characterize primary open angle glaucoma. Sci Rep. 2021;11(1):9766. doi:10.1038/s41598-021-89137-z
Ciotu IM, Stoian I, Gaman L, Popescu MV, Atanasiu V. Biochemical changes and treatment in glaucoma. J Med Life. 2015;8(1):28-31.
Ferreira SM, Lerner SF, Brunzini R, Evelson PA, Llesuy SF. Oxidative stress markers in aqueous humor of glaucoma patients. Am J Ophthalmol. 2004;137(1):62-69. doi:10.1016/s0002-9394(03)00788-8
Ghanem AA, Arafa LF, El-Baz A. Oxidative stress markers in patients with primary open-angle glaucoma. Curr Eye Res. 2010;35(4):295-301. doi:10.3109/02713680903548970
Benoist d'Azy C, Pereira B, Chiambaretta F, Dutheil F. Oxidative and Anti-Oxidative Stress Markers in Chronic Glaucoma: A Systematic Review and Meta-Analysis. PLoS One. 2016;11(12):e0166915. doi:10.1371/journal.pone.0166915
Goyal A, Srivastava A, Sihota R, Kaur J. Evaluation of oxidative stress markers in aqueous humor of primary open angle glaucoma and primary angle closure glaucoma patients. Curr Eye Res. 2014;39(8):823-829. doi:10.3109/02713683.2011.556299
Takayanagi Y, Takai Y, Kaidzu S, Tanito M. Evaluation of Redox Profiles of the Serum and Aqueous Humor in Patients with Primary Open-Angle Glaucoma and Exfoliation Glaucoma. Antioxidants (Basel). 2020;9(12):1305. doi:10.3390/antiox9121305
Liu A, Wang L, Feng Q, et al. Low expression of GSTP1 in the aqueous humour of patients with primary open-angle glaucoma. J Cell Mol Med. 2021;25(6):3063-3079. doi:10.1111/jcmm.16361
Cantó A, Olivar T, Romero FJ, Miranda M. Nitrosative Stress in Retinal Pathologies: Review. Antioxidants (Basel). 2019;8(11):543. doi:10.3390/antiox8110543
Bagnis A, Izzotti A, Centofanti M, Saccà SC. Aqueous humor oxidative stress proteomic levels in primary open angle glaucoma. Exp Eye Res. 2012;103:55-62. doi:10.1016/j.exer.2012.07.011